
Virus: Structure Types and Function
Structure of virus
Virion Structure is defined by capsid symmetry and presence or absence of an envelope. A complete virus particle is called a virion. Virion morphology has been intensely studied because of the importance of viruses and the realization that virion structure is simple enough to be understood in detail.
Progress has come from the use of several different techniques: electron microscopy, X-ray diffraction, biochemical analysis, and immunology. Although our knowledge is incomplete due to the vast diversity of viruses.
General Structural Properties of viruses
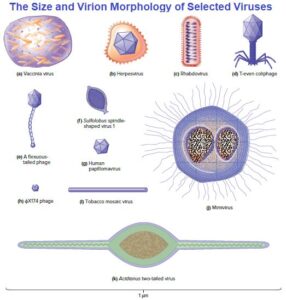
Virions can be extraordinarily tiny (about 20 nm in diameter) to about the same size as a rod-shaped bacterial cell (1.5 × 0.5 μm). The smallest are a little larger than ribosomes, whereas mimiviruses, among the largest viruses known, can be seen in the light microscope. However, most virus particles must be viewed with electron microscopes.
The simplest virions consist only of a nucleocapsid, which is composed of a nucleic acid, either DNA or RNA, and a protein coat called a capsid. The capsid surrounds the viral nucleic acid, protects the viral genome, and often aids in its transfer between host cells. Some virions are covered by a lipid membrane, and these are termed enveloped viruses, whereas those lacking a membrane are called non-enveloped or naked viruses.
Notice what is missing from viruses: Ribosomes for protein synthesis and a mechanism for generating ATP. Cytoplasm is absent, and while a few enzymes may be found, there are not enough to sustain cellular processes.
Non-enveloped viruses construct a capsid from many copies of one protein and a few minor proteins. Each subunit is termed a protomer, and thousands of protomers self-assemble to form the capsid. In contrast, enveloped viruses require both nucleocapsid proteins and additional proteins to anchor the membrane. Some viruses use noncapsid proteins as scaffolding upon which the capsids are assembled. Probably the most important advantage of this design strategy is that the viral genome is used with maximum efficiency.

For example, the tobacco mosaic virus (TMV) capsid is constructed using a single type of protomer. Recall that the building blocks of proteins are amino acids and that each amino acid is encoded by three nucleotides. The TMV protomer is 158 amino acids in length. Therefore only about 474 nucleotides are required to code for the coat protein. The entire TMV genome consists of only 6,400 nucleotides.
Thus only a small fraction of the genome is used to code for the capsid. Suppose, however, that the TMV capsid were composed of six different protomers, all about 150 amino acids in length. If this were the case, about 2,700 nucleotides in the TMV genome would be required just for capsid construction, and much less genetic material would be available for other purposes.
Helical Capsids of virus
Helical capsids are shaped like hollow tubes with protein walls. Tobacco mosaic virus is a well-studied example of helical capsid structurev. The self-assembly of TMV protomers into a helical arrangement produces a rigid tube. The capsid encloses an RNA genome, which is wound in a spiral and lies within a groove formed by the protein subunits. Not all helical capsids are as rigid as the TMV capsid. The nucleocapsids of influenza viruses are thin and flexible and are enclosed within an envelope.
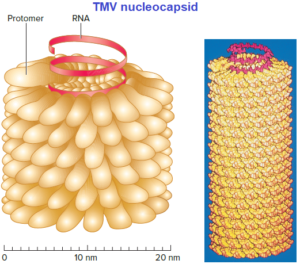
The size of a helical capsid is influenced by both its protomers and the viral genome. The diameter of the capsid is a function of the size, shape, and interactions of the protomers. The length of the capsid appears to be determined by the nucleic acid because a helical capsid does not extend much beyond the end of the viral genome.
Icosahedral Capsids of virus
An icosahedron is a regular polyhedron with 20 equilateral triangular faces and 12 vertices. Icosahedral capsids are the most efficient way to enclose a space. They are constructed from ring- or knob-shaped assemblages of five or six protomers; the assemblages are called capsomers.

Capsomers composed of five protomers are called pentamers (pentons); hexamers (hexons) are capsomers that possess six protomers. Pentamers are usually at the vertices of the icosahedron, whereas hexamers generally form its edges and triangular faces. The virions of some RNA viruses have pentamers and hexamers constructed with only one type of subunit. Other virions have pentamers and hexamers composed of different proteins. Although many icosahedral capsids contain both pentamers and hexamers, some have only pentamers.
Capsids of Complex Symmetry
Most viruses have either icosahedral or helical capsids, but some viruses do not fit into either category. Poxviruses and large bacteriophages are two important examples.

Poxvirus virions are among the largest of the animal viruses (about 400 by 240 by 200 nm in size) and can be seen with a light microscope. They possess an exceptionally complex internal structure with an ovoid- to brick-shaped exterior. The virion morphology of the vaccinia virus. Its double stranded DNA genome is associated with proteins and contained in the core, a central structure shaped like a biconcave disk and surrounded by a membrane. Two lateral bodies lie between the core and the virion’s outer envelope and contain viral enzymes.

Some large bacteriophages have virions that are even more elaborate than those of poxviruses. The virions of T2, T4, and T6 phages (T-even phages) that infect Escherichia coli are said to have binal symmetry because they have a head that resembles an icosahedron and a tail that is helical.
The icosahedral head is contains the DNA genome. The tail is composed of a collar joining it to the head, a central hollow tube, a sheath surrounding the tube, and a complex baseplate. In T-even phages, the baseplate is hexagonal and has a pin and a jointed long tail fiber and a short tail fiber at each corner.
Viral Envelopes and Enzymes
The nucleocapsids of many animal viruses, some plant viruses, and at least one bacterial virus are surrounded by an outer membranous layer called an envelope. Animal virus envelopes usually arise from the plasma or nuclear membranes of the host cell. Envelope lipids and carbohydrates are therefore acquired from the host. In contrast, envelope proteins are coded for by viral genes and may even project from the envelope surface as spikes, which are also called peplomers.
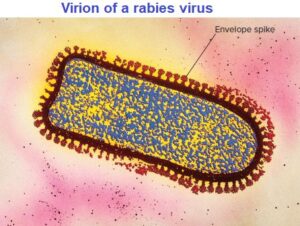
In many cases, spikes are involved in virion attachment to the host cell surface. Because spikes differ among viruses, they also can be used to identify some viruses. Many enveloped viruses have virions with a somewhat variable shape and are called pleomorphic. However, the envelopes of viruses such as the bullet-shaped rabies viruses are firmly attached to the underlying nucleocapsid and endow the virion with a constant, characteristic shape.
Influenza virus is a well-studied enveloped virus with two types of spikes. Some spikes consist of the enzyme neuraminidase, which functions in the release of mature virions from the host cell. Other spikes are hemagglutinin proteins, so named because they bind virions to red blood cells and cause the cells to clump together a process called hemagglutination.

Influenza virus’s hemagglutinins participate in virion attachment to host cells. Most of its envelope proteins are glycoproteins—proteins that have carbohydrate attached to them. A nonglycosylated protein, the M1 (matrix) protein, is found on the inner surface of the envelope and stabilizes both the lipid envelope and the display of both types of spike.
In addition to enzymes associated with the envelope or capsid (e.g., influenza neuraminidase), some viruses have enzymes within their capsids. Such enzymes are usually involved in nucleic acid replication. For example, influenza virus virions have an RNA genome and carry an enzyme that synthesizes RNA using an RNA template. Thus although viruses lack true metabolism and cannot reproduce independently of living cells, their virions may carry one or more enzymes essential to the completion of their life cycles.
Viral Genomes Are Structurally Diverse
One clear distinction between cellular organisms and viruses is the nature of their genomes. Cellular genomes are always doublestranded (ds) DNA. Viruses, on the other hand, employ all four possible nucleic acid types: dsDNA, single-stranded (ss) DNA, ssRNA, and dsRNA. All four types are used by animal viruses. Most plant viruses have ssRNA genomes, and most bacterial and archaeal viruses have dsDNA. The size of viral genomes also varies greatly.
Very small genomes are around 4,000 nucleotides—just large enough to code for three or four proteins. Some viruses save additional space by using overlapping genes. At the other extreme are the genomes of pandoraviruses,
which infect protists. They are about 2.5 × 106 nucleotides long, exceeding some bacteria and archaea in coding
capacity.
Most DNA viruses use dsDNA as their genetic material. However, some have ssDNA genomes (e.g., ϕX174 and M13). In both cases, the genomes may be either linear or circular. The ends of linear viral genomes may be covalently closed, attached to a protein, or otherwise masked. Some DNA genomes can switch from one form to the other. For instance, the E. coli phages lambda has a linear genome in its capsid, but it becomes circular once it enters the host cell.
Relatively few RNA viruses have dsRNA genomes. More common are viruses with ssRNA genomes. Polio, tobacco mosaic, rabies, influenza, and human immunodeficiency (HIV) are all ss-RNA viruses.
Some RNA viruses have segmented genomes—genomes that consist of multiple pieces (segments) of RNA. In many cases, each segment codes for one protein and there may be as many as 10 to 12 segments. Usually all segments are enclosed in the same capsid. However, the genome of brome mosaic virus, a virus that infects certain grasses, is composed of three segments distributed among three different virions.
Reference and Sources
- https://www.slideshare.net/PRIYAKUMARI45/structural-symmetry-of-viruses
- https://www.researchgate.net/publication/233394888_Assembly_stability_and_dynamics_of_virus_capsids
- https://quizlet.com/149231652/prescott-microbiology-chapter-6-flash-cards/
- https://www.researchgate.net/publication/282127373_The_rapidly_expanding_universe_of_giant_viruses_Mimivirus_Pandoravirus_Pithovirus_and_Mollivirus
- http://www.science.gov/topicpages/t/teno+sus+viruses.html
- https://www.researchgate.net/publication/276146730_Influenza_virusmediated_membrane_fusion_Structural_insights_from_electron_microscopy
- https://quizlet.com/170615486/prescott-microbiology-chapter-6-viruses-final-flash-cards/
- https://www.gscgandhinagar.in/assets/admin/images/Unit%201General%20characteristics%20of%20viruses.pdf
- https://en-academic.com/dic.nsf/enwiki/11126443
- https://quizlet.com/20062151/microbiology-flash-cards/
- https://quizlet.com/ca/277067578/biol232-midterm-2-flash-cards/
- https://www.researchgate.net/profile/Tetsuro-Okuno/publication/15604530_Inhibition_of_brome_mosaic_virus_BMV_amplification_in_protoplasts_from_transgenic_plants_expressing_replicable_BMV_RNAs/links/0912f5112f4772da09000000/Inhibition-of-brome-mosaic-virus-BMVamplification-in-protoplasts-from-transgenic-plants-expressing-replicable-BMV-RNAs.pdf
Also Read:
- SARS: Evolution of a Virus
- what is microbiology?
- AIDS: Acquired Immune Deficiency Syndrome
- Classification of viruses on the basis of genome
- Virus: Introduction, Properties and Classifications
- Cas9 Nuclease for Genome Editing
- Classification of viruses on the basis of genome
- Fundamental Principle of Clinical Specimen Collection
- Reverse Transcription Polymerase Chain Reaction (RT-PCR)
- Viruses Arise from Genetic Recombination and Mutation
- Viruses associated with Human Tumors and Cancers
- Tools of Genetic Engineering to Make Genome Modifications
- Malaria: Causative Agent, Symptoms, Treatment and Prevention
- Comparison Between the Domains Bacteria, Archaea, and Eukarya
- Hansen’s Disease: Symptoms, Causative Agent, Pathogenesis and Treatment
- Gonorrhea: Causative Agent, Symptoms, Treatment and Prevention