The nucleus is a central part of the cell; it can be easily distinguished between prokaryotic and eukaryotic cells.
The nucleus functions as a database of the cell genome; it controls the genomic expressions of the cell.
In fact, the nucleus is responsible for providing a site for the process of replication and transcription of the DNA, even RNA is processed in the nucleus before the translation mechanism, which usually occurs in the cytoplasm.
The nucleus consists of the nuclear envelope which includes nuclear pores, nuclear lamina, in the center of the nucleus, the nucleolus is present, which is surrounded by dense chromatin.
In prokaryotes, instead of a well-structured nucleus, nucleoid is present, therefore steps of central dogma are altogether regulated, which means consecutively translation is regulated, while transcription is going on.
The nuclear envelope performs several roles in the functioning of the nucleus exists in the eukaryotes.
It separates the genome from the cytoplasmic surrounding.
It limits the access of selected proteins entering the area of genetic material i.e., exists in the center of the nucleus.
It controls the genomic expression of the mRNA because transcription factors are moved from the cytoplasm to the nucleus, through a nuclear pore. The whole process is well regulated.
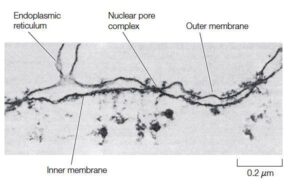
Electron micrographic image of the Nucleus
Detailed Structure of the Nucleus
The nuclear envelope serves as a framework structure of the nucleus. It restricts the molecules to move across the nucleus membrane, forming a distinct compartment.
The exchange of the selected molecules including co-factors, proteins are regulated by nuclear pore complexes, gene expression regulation is well maintained.
It constitutes two nuclear membranes, nuclear pore complexes & nuclear lamina.
Nuclear Membranes
Nuclear membranes are composed of a phospholipid bilayer that surrounds the nucleus and is called inner & outer nuclear membranes.
Usually, the endoplasmic reticulum [ER] is connected to the nucleus via external nuclear membranes that are an outermost region of the nucleus. In fact, external membranes are the extension of the ER, and space between two nuclear membranes is directly connected to the lumen of the ER.
Ribosomes bounds to the cytosolic surrounding of the external nuclear membranes, which are enriched with membrane proteins, which initiate binding with the cytoskeleton.
The inner nuclear membrane link with the nuclear lamina followed by specific integral membranes proteins about 60 in numbers.
Image showing outer and inner nuclear membranes
Nuclear Pore complex
Both nuclear membranes joined at pore complexes of the nucleus. It is known as channels that provide passage to the specific molecules and ions generally polar in nature and macromolecules [Proteins & RNA] that are needed for gene regulation.
The nuclear pore complex is an approximate molecular weight of about 120 million Da.
The nuclear pore complex consists of several copies of pore proteins about 30 in number which is also termed nucleoporins or NUPs.
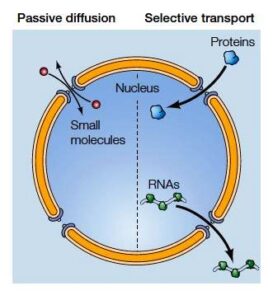
Transfer of molecules by nuclear pore complex is carried out by two mechanisms
Small molecules with less molecular mass up to 40kd, moves freely across the pore complex by diffusion.
Most macromolecules like RNA & Proteins use selective transport systems for their movement across the pore complex. Generally, selective transport is triggered in response to the specific signals which regulate the transport.
It works as a boundary that isolates the nucleus region from the cytoplasm and also restricts the mobility of the molecules crossing the nuclear membrane.
Types of molecular movement through nuclear pore complexes
The basic task of the pore complex is to control the traffic of the specified molecules across the nucleus, also manage the transfer of the proteins which is significant for gene regulation, for instance, transcription factors synthesize in the cytoplasm & transported to the nucleus for their contributing role in transcription.
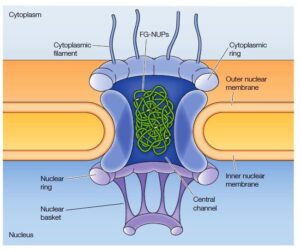
Structure of Nuclear Pore Complex
Nuclear Pore complex
The 3-D structural information of the nuclear pore complex is revealed through electron microscopy.
It constitutes the eight spokes assembled in the interior of the channel, which is connected with the cytoplasmic rings and the complete channel is anchored inside the nuclear envelope particularly at the intersection of the fused nuclear membranes [i.e., outer & inner membranes fused at the point].
The extended nuclear pore complex represents the basket-like shape.
The central section of the channel consists of proteins known as FG-NUPs, rich in repetitive glycine and phenylalanine amino residues, it serves as a barrier that determines the nuclear pore permeability and regulates the selective transport.
Nuclear Lamina
It is usually found beneath the inner nuclear membrane, composed of filamentous meshwork, which structurally supports the nucleus.
The nuclear lamina constitutes fibrous proteins termed as lamins, about 60- to 80 kDa mol mass, along with different proteins.
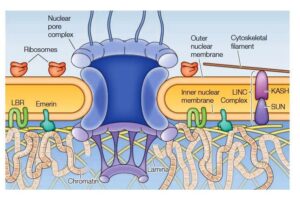
Image showing structural detail of the nuclear lamina
Lamins are related to the class of filament proteins which form a linkage between each other in a very highly arranged manner.
Initially, two lamins proteins form a dimer, in which two polypeptide α-helical regions intertwined themselves in a structure, termed as coiled-coil, again lamin dimers form linkage for building the filaments, which is the foundation of the nuclear lamina.
Additionally, the inner nuclear membrane associates itself with the lamins filaments, which is accompanied by the addition of the lipids, particularly prenylation of cysteine residues at the C-terminal.
The inner nuclear membrane has specific proteins which include emerin & lamin- B receptors, which binds to the lamins proteins.
The nuclear lamina association is completed by connecting themselves to the protein complexes i.e., the LINC complex exists in the cytoskeleton which spans the nuclear membranes.
It is also observed through studies lamin and its associated protein attached to the chromatin, exist in the core of the nucleus.
Nucleolus and Chromatin
Gene is highly organized in the chromatin structure, dense in appearance during the cell cycle, it is visually prominent through the microscopic view.
Generally, two types of chromatin exist in the core of the nucleus, one is Euchromatin, which is not condensed and transcriptionally active, on the other hand, heterochromatin, is highly packed and condensed, not active for transcription.
Nucleolus serves as a site of transcription, processing of rRNA, and also assembly of ribosomes.