The biological membranes structure is made up of lipids that exist in the bilayer, separating cytosol from the extracellular environment with the help of a defined boundary.
The membrane bilayer constitutes 50% of each lipid and protein by mass, it also includes a carbohydrate group associated with a lipid called glycolipid or associated with a protein called glycoprotein about 5-10% of membrane mass.
Maximum of the vital tasks are executed by membrane proteins placed in a bilayer, which provides different functional properties to the individual cell’s plasma membranes.
The proteins present in membranes, their types and amounts usually differ in individual cell types.
For instance, myelin membrane present in axons of nerve cells serves as the insulating layer which helps electrical impulses to travel far in the nerve cells, constitute 25% protein of its membrane mass, by contrast, membrane bilayer of organelle mitochondria & chloroplasts involves in the manufacture of ATP, constitutes approximately 75% of protein.
Model Representation of Membrane Organization in the Structure
In 1972, Jonathan Singer and Garth Nicolson suggested the membrane model structure as fluid bilayer, in which different types of proteins present in the membrane are embedded, and both lipids along with membrane proteins can diffuse sideways lateral in the membrane’s plane.
Schematic description of Structure of membrane showing various membrane proteins embedded in bilayer
Proteins present in the membranes diffuses in the fluidic bilayer
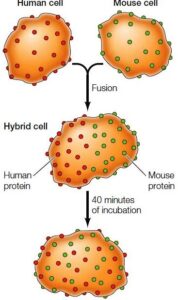
In 1970, an experiment was carried out by Larry Frye and Michael Edidin which determined the membrane protein’s diffusion.
In their experiment, two cells from different origins produced hybrid cells by fusion (by using human and mouse cells), on the hybrids, the proteins were distributed on membranes which were analyzed by using specific fluorescent-labeled antibodies to identify proteins of mouse & human origin.
Within 40 minutes, mouse and also human membrane proteins were merged on the hybrid cell surface, which shows proteins moved freely through the fluidic bilayer.
The experiment concludes diffusion of proteins within the membrane’s plane
The fluidity of bilayer and its proteins that exist in the membranes is depicted by the model sets a basic example of how the different membrane proteins, as well as lipids, organize themselves within membranes of all cell types.
Two Major Classes of Membrane- Associated Proteins
The proteins present in membranes were classified into two major classes by Singer and Nicolson, which were known as peripheral & integral membrane proteins.
The peripheral membrane proteins are indirectly linked with the bilayer and avoid the interaction of the hydrophobic group within the membrane.
By treating with reagents, mostly polar in nature like solutions of high concentration of salt or extremely high pH, proteins dissociate themselves without disrupting the membrane bilayer.
The integral membrane proteins, fixed inside the membrane bilayer, associate themselves with the hydrophobic group of the membranes; therefore they can only be dissociated by reagent which disrupts the hydrophobic interactions of the phospholipid bilayer.
The most common reagents, which are used to disrupt the hydrophobic groups interacting with the bilayer, are detergents.
Detergents contain both hydrophilic and hydrophobic groups and are amphipathic in nature. The hydrophobic domains of detergents replace the lipids membrane and linked themselves with the interior side of the integral membrane. The complexes which are formed are dispersable in aqueous solutions since the detergent molecule on the opposite side is hydrophilic in nature.
Detergents solubilizing proteins of the membrane
Nearly all of the integral proteins are considered transmembrane proteins, which are amphiphilic in nature, which have both hydrophilic and hydrophobic regions.
The hydrophobic domain passes through the membrane bilayer and covalently interacts with the interior of the bilayer which consists of a hydrophobic group, and the other end is hydrophilic which is exposed on the opposite side of the membranes bilayer.
In the membrane bilayer, inserted proteins have a special orientation, are arranged asymmetrically, and have individual functions of both cytosolic and non-cytosolic regions of the domains.
Both domains are divided by, the transmembrane proteins segment of the polypeptide chain spanning the membrane, which largely contains non-polar side chains amino acids, because peptide bonds are polar, so all peptides bonds in bilayer create H- bonds with each another in scarcity of water, in α- helix form, maximum hydrogen bonds are formed between peptide bonds, by transversing segment of transmembrane proteins spanning the membrane bilayer.
A membrane-spanning segment of transmembrane protein as an α-helix
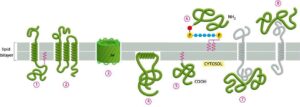
Membrane proteins interact with the membrane in many ways
The membrane linked with proteins in a bilayer in the following manner is shown below figure.
In different ways , membrane proteins associate with the lipid bilayer.
Several transmembrane proteins extend themselves, crossing the bilayer which exists in α-helix form, single-pass is covalently attached with fatty acid situated in the inner monolayer. e.g. Bacteriorhodopsin
Some are multi-pass transmembrane proteins, which exist in multiple α-helix forms.
In a rolled-up form b- sheet (called b- barrel), e.g., aquaporins, which allow passage of polar molecules.
Few proteins of membranes exist in the cytosol and form interactions with its monolayer, by amphiphilic α-helix.
Some proteins present in the membrane, fatty acid or prenyl group are covalently attached with membrane bilayer.
Other proteins link with the membrane are attached with oligosaccharide linker to phosphatidylinositol, within monolayer of non- cytosolic side called glycosylphosphatidylinositol (GPI) anchor.
Many proteins bind with other membrane proteins by non-covalent.
Membrane proteins link with the cytoskeleton and restrict Membrane Proteins Diffusion
The erythrocyte is used as an example to study the structure of membrane and membrane proteins. The human erythrocytes have various proteins link with membranes and originally identification is carried out by gel electrophoresis. Several proteins exist in the periphery of the plasma membrane termed, as peripheral membrane proteins, which form an association with cortisol cytoskeleton which is present beneath the membrane.
The cell restricts the motion of the specific proteins present in the membrane by tethering them to the bilayer.
Four ways to restrict movement of membrane proteins
Large aggregates tethered by macromolecule interaction e.g bacteriorhodopsin.
Interaction of macromolecule on the exterior the cell,
Interact within the bilayer
Interact with the protein exist on the exterior surface of another cell.
The richest peripheral protein of RBC present on its membrane is spectrin, other proteins like actin, ankyrin, & band 4.1 are also present. Membrane associate itself with cytoskeleton with the help of protein ankyrin, it serves to bind together spectrin and band 3.1 protein. Additionally, band 4.1 provides a link between membrane and cytoskeleton, which hold up glycophorin, which is a crucial membrane protein of RBC.
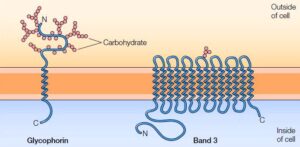
The two dominating classes of membrane proteins also called integral membrane proteins of erythrocytes are Glycophorin & band 3.
Glycophorin, a small glycoprotein composed of the amino groups about 131 residues (molecular weight about 30,000) which traverses the bilayer with a single spanning segment, an α- helix made of the amino group around 23 in number. The polypeptide chains containing amino residues are densely glycosylated which is revealed on the exterior of the cell membrane.
Band 3.1, work as a transporter that carries anions, which includes Chloride (Cl–) and bicarbonate (HCO3). It is composed of amino groups of around 929 residues with 14 membrane-spanning α-helices. Band 3 forms dimer inside the membrane, form a globular structure that contains channel protein which serves passage for the ions crossing the lipid bilayer.
Integral membrane proteins of Erytrocyte- Glycophorin and Band 3
Membrane Proteins are related to the carbohydrates group
Several transmembrane proteins, which are densely glycosylated, due to which, the carbohydrates residue is found on the exterior cell surface membrane interacting with glycolipid or glycoprotein.
The surface of the cell is packed with a carbohydrates coat called glycocalyx, which is made by oligosaccharides present in both lipids & proteins.
It serves as a protective barrier against ionic and mechanical stress.
Electron Microscopic image of glycocalyx of epithelial cells of the intestine
Hydropathy Plot
The membrane protein, like glycophorin, constitutes of 20-25 amino group in their α- helical region, which spans bilayer about the thickness of 30 Å.
Several amino groups in the spanning region are non-polar and uncharged.
This information is used in the identification of spanning regions in a cell membrane protein.
Once the amino acid sequence of the protein’s spanning region is known, the number and positions of transmembrane protein could be obtained from a hydropathy plot, also termed as hydrophobicity plot.
This method is used to find out the nature of regions that might be hydrophilic or hydrophobic exist in the protein sequence.
It is estimated through the hydropathy plot, 20% of organism’s proteins are transmembrane proteins.
Hydropathy plots, locate possible α -helical membrane-spanning segments A. Glycophorin, one peak resembles single- transmembrane spanning segment B. Bacteriorhodopsin, seven peaks resemble seven-transmembrane spanning segment
Experimentally, the polarity of individual amino groups can be concluded by calculating the alteration in their free energy followed by the movement of that amino group’s side chain from hydrophobic solvent to water. The alteration in free energy is inferred by the hydropathy index.
A positive value on the hydropathy index indicates the free energy needed for transfer to water, which means the segment is hydrophobic; the value mentioned is an index of the amount of energy needed.
The combined hydropathy index of a sequence can be inferred by adding the free energies transfer for the amino group in the sequence.
To evaluate a polypeptide sequence for membrane-spanning segments, hydropathy index is calculated for consecutive segments of given size about 10-20 amino acids.