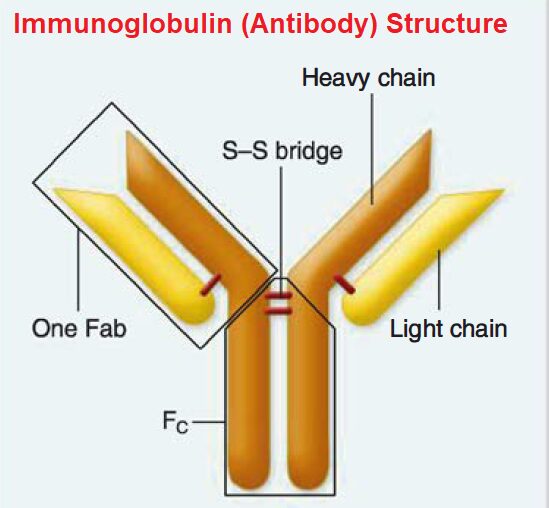
All immunoglobulin molecules have a basic structure composed of four polypeptide chains.
Two identical heavy chains.
Two identical light chains.
A computer generated model of case, antibody specificity results from the nature of antibody-antigen binding. Immunoglobulin structure showing the arrangement of the four polypeptide chains.
Light-chain polypeptide mainly consists of 220 amino acids and has a mass of 25,000 Da.
Each heavy chain consists of around 440 amino acids and has a mass of 50,000 to 70,000 Da.
Heavy and light chains are connected to each other by disulfide bridges or bonds.
The heavy chains are structurally distinct for each immunoglobulin class or subclass.
Both light (L) and heavy (H) chains contain two different regions.
The constant (C) regions (CL and CH) have amino acid sequences that do not vary significantly between antibodies of the same class.
The variable (V) regions (VL and VH) have different amino acid sequences, and these regions fold together to form the antigen-binding sites.
Heavy and light chains are connected to each other by disulfide bridges (bonds). Each heavy and light chain pair form an antigen-binding fragment (Fa b). The disulfide-linked heavy chains form the crystalizable fragment (Fe).
The four chains are in the shape of a Y with a hinge region. This hinge provides the antibody molecule to be more flexible, adjusting to the different arrangements of the epitopes on antigens.
The stalk of the Y is termed the crystallizable fragment (Fe) and can bind to a host cell by interacting with the cell-surface Fe receptor.
The top of the Y consists of two antigen- binding fragments (Fab) that bind with compatible epitopes.
The Fe fragments are composed only of constant regions, whereas the Fab fragments have both constant and variable regions.
Both the heavy and light chains contain several homologous units of about 100 to 110 amino acids.
Within each unit, called a domain, disulfide bonds form a loop of approximately 60 amino acids.
Interchain disulfide bonds also link heavy and light chains together.
All light chains contain a single variable domain (VL) and a single constant domain (CJ. Heavy chains contain a variable domain (VH) and either three or four constant domains (CH1, CH2, CH3, and CH4). The variable regions (VH, VL), when folded together in three-dimensions, form the antigen-binding sites.
The light chain may be either of two distinct forms called kappa (K) and lambda (A).
These can be distinguished by the amino acid sequence of the constant (C) portion of the chain.
In humans, the constant regions of all chains are identical. Each antibody molecule produced by a sole B cell will contain either K or A light chains but never both.
The light-chain variable (V) domain contains hyper variable regions, or complementarity determining regions (CDRs), that differ in amino acid sequence more frequently than the rest of the variable domain.
The heavy chain also has a variable region (VH) at its amino terminal domain.
The other domains of the heavy chains are termed constant (C) domains.
The constant domains of the heavy chain form the constant (CH) region. The amino acid sequence of this region determines the classes of heavy chains.
In humans, there are five different classes of heavy chains designated by lowercase Greek letters: gamma (γ), alpha (α) , mu (μ), delta (δ), and epsilon (ε), and usually written as G, A, M, D, or E.
The properties of these heavy chains determine, respectively, the five immunoglobulin (Ig) classes: IgG, IgA, IgM, IgD, and IgE.
Each immunoglobulin class differs in its general properties, half-life, distribution in the body, and interaction with other components of the host’s defensive systems.
Immunoglobulin Function
Each end of the immunoglobulin molecule has a unique role.
The Fab region is concerned with binding antigen, whereas the Fe region mediates binding to receptors (called Fe receptors) found on various immunocytes or the first component of the classical complement system.
The antibody serves to mark and identify the non-self agent as a target for immunological attack and to activate innate immune responses that can destroy the target.
An antigen binds to an antibody at the antigen-binding site within the Fab region of the antibody.
More specifically, a pocket is formed by the folding of the VH and VL regions.
At this site, specific amino acids contact the antigen’s epitope and form multiple noncovalent bonds between the antigen and amino acids of the binding site.
Because binding is due to weak, noncovalent bonds such as hydrogen bonds and electrostatic attractions, the antigen’s shape must exactly match that of the antigen-binding site.
If the shapes of the epitope and binding site are not truly complementary, the antibody will not effectively bind the antigen.
Thus a lock-and-key mechanism operates; however, in at least one case, the antigen binding site changes shape when it complexes with the antigen (an induced-fit mechanism).
In either case, antibody specificity results from the nature of antibody-antigen binding.
Phagocytes have Fe receptors for immunoglobulin on their surface, so bacteria that have antibodies are targets for phagocytosis by neutrophils and macrophages, this is called as opsonization.
Other cells, like natural killer cells, destroy antibody-coated cells by antibody-dependent cell-mediated cytotoxicity.
Immune destruction also is promoted by antibody-induced activation of the classical complement system.